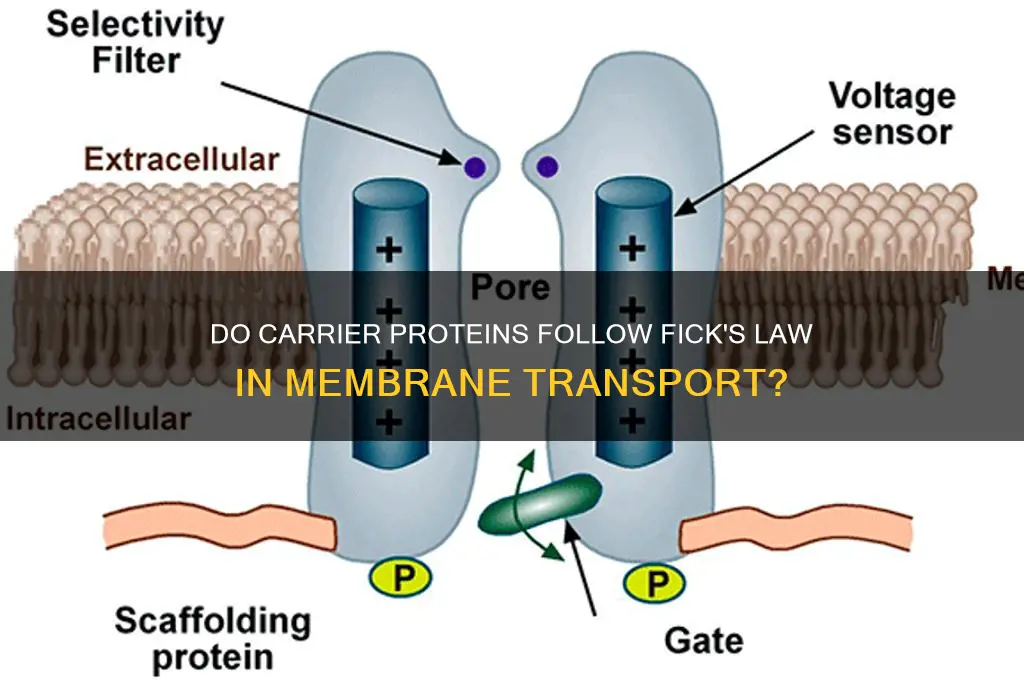
Carrier proteins, which facilitate the transport of molecules across biological membranes, present an intriguing question regarding their adherence to Fick's laws of diffusion. Fick's laws describe the passive movement of particles from regions of higher concentration to lower concentration, driven by a concentration gradient. While carrier-mediated transport involves protein-substrate interactions and often requires energy, it is still influenced by concentration differences. The relationship between carrier proteins and Fick's laws is complex, as these proteins can exhibit both passive and active transport mechanisms. In passive transport, carrier proteins may follow Fick's laws, allowing molecules to move down their concentration gradient. However, in active transport, carrier proteins can move molecules against their concentration gradient, seemingly defying Fick's laws. Understanding how carrier proteins operate within the framework of Fick's laws is crucial for comprehending the dynamics of molecular transport in biological systems.
| Characteristics | Values |
|---|---|
| Obedience to Fick's Law | Carrier proteins do not strictly obey Fick's Law of diffusion. Fick's Law describes passive diffusion of solutes across a concentration gradient, which assumes no interaction with the membrane. Carrier proteins, however, facilitate active or facilitated transport, often against concentration gradients, involving specific binding and conformational changes. |
| Transport Mechanism | Carrier proteins use facilitated diffusion or active transport, depending on the energy source. Facilitated diffusion is passive (down a gradient), while active transport requires energy (ATP or ion gradients). |
| Specificity | Highly specific to particular molecules or ions, unlike simple diffusion through lipid bilayers. |
| Saturation | Transport rates saturate at high substrate concentrations due to limited carrier protein availability, unlike linear diffusion rates in Fick's Law. |
| Inhibition | Can be inhibited by specific inhibitors or competitive substrates, unlike passive diffusion. |
| Energy Dependence | Some carrier proteins (e.g., in active transport) require energy, while Fick's Law describes energy-independent processes. |
| Directionality | Can transport molecules against their concentration gradient (active transport), which violates the passive nature of Fick's Law. |
| Regulation | Transport can be regulated by factors like pH, temperature, or cellular signals, unlike simple diffusion. |
| Examples | GLUT transporters (glucose), Na+/K+ ATPase (active transport), and aquaporins (water channels). |
| Mathematical Description | Transport by carrier proteins is often described by Michaelis-Menten kinetics, not Fick's Law. |
Explore related products






What You'll Learn
- Passive vs. Active Transport: Does Fick's Law Apply to Both Mechanisms
- Role of Concentration Gradients in Carrier Protein-Mediated Transport
- Impact of Saturation on Fick's Law in Carrier Protein Systems
- How Protein Specificity Influences Fick's Law in Membrane Transport?
- Experimental Evidence: Testing Fick's Law in Carrier Protein Dynamics
![]()
Passive vs. Active Transport: Does Fick's Law Apply to Both Mechanisms?
Fick's Law, a fundamental principle in diffusion, describes how particles move from an area of high concentration to an area of low concentration across a concentration gradient. It is a cornerstone in understanding passive transport, a process where substances move across cell membranes without requiring energy input from the cell. In passive transport, molecules such as oxygen, carbon dioxide, and lipids diffuse through the lipid bilayer of the cell membrane, following the principles outlined by Fick's Law. This mechanism is essential for maintaining cellular homeostasis and ensuring that cells receive necessary nutrients and expel waste products efficiently.
When considering carrier proteins, which facilitate the movement of specific molecules across the cell membrane, the application of Fick's Law becomes more nuanced. Carrier proteins are involved in both passive (facilitated diffusion) and active transport. In facilitated diffusion, carrier proteins assist in the movement of molecules down their concentration gradient, a process that still aligns with Fick's Law. Here, the carrier proteins act as channels or carriers that increase the rate of diffusion without altering the direction, which is dictated by the concentration gradient. Thus, in passive transport involving carrier proteins, Fick's Law remains applicable, as the movement is driven by the natural tendency of molecules to disperse.
In contrast, active transport involves the movement of molecules against their concentration gradient, from an area of low concentration to an area of high concentration, requiring energy (often in the form of ATP). This process directly opposes the principles of Fick's Law, as it does not rely on the natural diffusion of molecules. Carrier proteins in active transport, such as the sodium-potassium pump, use energy to transport molecules uphill, defying the concentration gradient. Therefore, Fick's Law does not apply to active transport mechanisms, as the movement is not driven by diffusion but by energy-dependent processes.
The distinction between passive and active transport highlights the limitations of Fick's Law in describing all forms of molecular movement across membranes. While Fick's Law is a powerful tool for understanding passive transport, including facilitated diffusion mediated by carrier proteins, it falls short in explaining active transport, where energy plays a dominant role. This difference underscores the complexity of membrane transport mechanisms and the need to consider both thermodynamic principles (like Fick's Law) and bioenergetic processes when studying cellular transport.
In summary, Fick's Law applies to passive transport mechanisms, including facilitated diffusion involving carrier proteins, as these processes rely on the concentration gradient for molecular movement. However, active transport, which uses energy to move molecules against their gradient, does not obey Fick's Law. Understanding this distinction is crucial for comprehending how carrier proteins function in different transport mechanisms and how cells regulate the movement of substances across their membranes. By integrating knowledge of Fick's Law with the energetics of active transport, researchers can gain a more comprehensive view of membrane transport dynamics.
Understanding UK Law: What Constitutes Threatening Behaviour?
You may want to see also
Explore related products






![]()
Role of Concentration Gradients in Carrier Protein-Mediated Transport
Carrier protein-mediated transport is a fundamental process in biology, facilitating the movement of specific molecules across cell membranes. This mechanism is crucial for maintaining cellular homeostasis, nutrient uptake, and waste removal. At the heart of this process lies the concept of concentration gradients, which play a pivotal role in driving the transport of molecules. Concentration gradients refer to the difference in the concentration of a substance across a membrane, from an area of higher concentration to an area of lower concentration. In the context of carrier proteins, these gradients act as the driving force for both facilitated diffusion and active transport.
Carrier proteins, also known as permeases or transporters, are transmembrane proteins that bind specific molecules and facilitate their movement across the membrane. Unlike simple diffusion, which relies solely on the kinetic energy of molecules, carrier protein-mediated transport is highly selective and regulated. The question of whether carrier proteins obey Fick's Law—a principle describing the diffusion of particles from higher to lower concentration—is nuanced. While Fick's Law accurately describes the direction of movement, carrier protein-mediated transport involves additional complexities, such as protein binding kinetics and conformational changes, which are not accounted for in Fick's Law. However, the underlying principle of concentration gradients remains central to the process.
In facilitated diffusion, carrier proteins transport molecules down their concentration gradient, requiring no additional energy input. The concentration gradient provides the thermodynamic driving force, as molecules move from an area of higher concentration outside the cell to an area of lower concentration inside, or vice versa. This process is passive and saturable, meaning it reaches a maximum rate when all carrier proteins are occupied. The role of the concentration gradient here is straightforward: it determines the direction and rate of transport, with steeper gradients leading to faster movement of molecules.
In contrast, active transport involves carrier proteins moving molecules against their concentration gradient, from an area of lower concentration to an area of higher concentration. This process requires energy, often in the form of ATP, to drive the transport. Despite the energy expenditure, concentration gradients still play a critical role. The gradient acts as a "potential difference" that the carrier protein works against, ensuring the molecule is transported to where it is needed. For example, the sodium-potassium pump actively transports sodium ions out of the cell and potassium ions into the cell, maintaining steep concentration gradients essential for cellular functions like nerve impulse transmission.
The interplay between concentration gradients and carrier proteins is further highlighted in secondary active transport, where the energy from one concentration gradient (e.g., sodium ions) is used to drive the transport of another molecule against its gradient. This mechanism underscores the importance of concentration gradients in both direct and indirect ways. While carrier proteins do not strictly obey Fick's Law due to their complex mechanisms, the principles of concentration gradients remain indispensable for understanding their function.
In summary, concentration gradients are the cornerstone of carrier protein-mediated transport, dictating the direction and efficiency of molecular movement across membranes. Whether in facilitated diffusion, active transport, or secondary active transport, these gradients provide the necessary driving force for carrier proteins to function. While Fick's Law provides a foundational framework for understanding diffusion, the role of concentration gradients in carrier protein-mediated transport is far more intricate, involving protein dynamics and energy considerations. Thus, concentration gradients are not just passive facilitators but active determinants of cellular transport processes.
Minnesota Bike Helmet Laws: What You Need to Know
You may want to see also
Explore related products


![]()
Impact of Saturation on Fick's Law in Carrier Protein Systems
Fick's laws of diffusion describe how particles spread out in a medium due to random motion, driven by a concentration gradient. In biological systems, carrier proteins facilitate the transport of specific molecules across membranes, often against their concentration gradient. A critical question arises: do carrier proteins obey Fick's law? The answer is nuanced, especially when considering the impact of saturation on these systems. While carrier proteins do not strictly follow Fick's laws due to their active transport mechanisms, understanding how saturation influences their behavior provides valuable insights into their transport kinetics.
Saturation occurs when all available binding sites on carrier proteins are occupied by the substrate molecules. At this point, further increases in substrate concentration do not lead to a proportional increase in transport rate. This phenomenon significantly impacts the applicability of Fick's laws in carrier protein systems. Fick's first law assumes a linear relationship between concentration gradient and flux, which holds true for simple diffusion. However, in carrier-mediated transport, the flux approaches a maximum (Vmax) as saturation is reached, deviating from linearity. This nonlinear behavior highlights the limitations of Fick's laws in describing carrier protein transport under saturating conditions.
The impact of saturation becomes more pronounced when analyzing Fick's second law, which describes how concentration changes over time. In unsaturated conditions, carrier proteins can maintain a steady flux proportional to the concentration gradient, somewhat aligning with Fickian diffusion. However, as saturation is approached, the transport rate plateaus, leading to a concentration profile that deviates from the predictions of Fick's second law. This deviation is particularly evident in systems with high substrate concentrations, where the transport rate is limited by the number of carrier proteins rather than the concentration gradient.
To model carrier protein systems under saturation, alternative approaches such as Michaelis-Menten kinetics are more appropriate. The Michaelis-Menten equation accounts for the saturation effect by incorporating Vmax and the Michaelis constant (Km), which represents the substrate concentration at half-maximal velocity. This framework provides a more accurate description of transport kinetics in carrier protein systems, especially when saturation is a dominant factor. By integrating these kinetic principles, researchers can better understand how carrier proteins operate in biological contexts, where substrate concentrations often fluctuate and may reach saturating levels.
In conclusion, while carrier proteins do not strictly obey Fick's laws, the impact of saturation on their transport behavior is a critical aspect to consider. Saturation introduces nonlinearity and limits the applicability of Fickian diffusion principles, necessitating the use of alternative models like Michaelis-Menten kinetics. Understanding these dynamics is essential for studying membrane transport processes, drug delivery systems, and metabolic pathways, where carrier proteins play a central role. By acknowledging the limitations of Fick's laws in saturated carrier protein systems, researchers can develop more accurate and predictive models of molecular transport in biological systems.
Understanding Michigan's Tint Law: Rules, Regulations, and Legal Limits
You may want to see also
![]()
How Protein Specificity Influences Fick's Law in Membrane Transport
Fick's laws of diffusion describe how particles spread within a medium due to random motion, driven by a concentration gradient. In the context of membrane transport, Fick's laws are often applied to passive diffusion across biological membranes. However, the presence of carrier proteins, which facilitate the movement of specific molecules across membranes, introduces a layer of complexity. Carrier proteins exhibit specificity, meaning they selectively bind and transport particular substrates. This specificity significantly influences how Fick's laws operate in membrane transport, as it deviates from the simple, concentration-driven diffusion described by Fick's equations.
Carrier proteins operate through facilitated diffusion or active transport, depending on whether the transport is down or against the concentration gradient, respectively. In facilitated diffusion, carrier proteins still obey Fick's laws in principle, as the direction and rate of transport are determined by the concentration gradient of the substrate. However, the specificity of the carrier protein restricts transport to only those molecules that can bind to the protein. This means that even if a concentration gradient exists for a non-specific molecule, it will not be transported, violating the assumption of Fick's laws that all particles diffuse equally. Thus, while the overall process still aligns with Fick's principles, the specificity of the carrier protein acts as a filter, limiting transport to specific substrates.
In active transport, carrier proteins use energy (e.g., ATP) to move molecules against their concentration gradient. Here, Fick's laws are less directly applicable because the transport is not solely driven by diffusion. Instead, the specificity of the carrier protein determines which molecules are actively transported, regardless of the concentration gradient. This process introduces a non-Fickian element, as the transport rate is influenced by the protein's binding affinity, energy availability, and conformational changes, rather than just the concentration difference. Therefore, while Fick's laws provide a foundational framework, the specificity of carrier proteins in active transport necessitates a more nuanced understanding of membrane transport dynamics.
The binding affinity and saturation of carrier proteins further modulate their influence on Fick's laws. High binding affinity ensures that only specific molecules are transported efficiently, while saturation limits the transport rate even if the concentration gradient persists. This behavior contrasts with simple diffusion, where the transport rate is directly proportional to the concentration gradient. Thus, carrier protein specificity introduces a regulatory mechanism that Fick's laws alone cannot account for, as they assume unrestricted diffusion of all particles.
In summary, carrier proteins do not strictly obey Fick's laws in membrane transport due to their specificity. While facilitated diffusion retains some alignment with Fick's principles, active transport and the selective nature of carrier proteins introduce deviations. The specificity of these proteins acts as a critical determinant of transport efficiency, substrate selectivity, and directional movement, making Fick's laws an incomplete descriptor of membrane transport in the presence of carrier proteins. Understanding this interplay is essential for modeling and predicting transport phenomena in biological systems.
Wesley's Legal Woes: License Loss?
You may want to see also
![]()
Experimental Evidence: Testing Fick's Law in Carrier Protein Dynamics
The question of whether carrier proteins obey Fick's laws of diffusion is a nuanced one, and experimental evidence plays a crucial role in addressing this. Fick's laws describe the passive diffusion of solutes from regions of high concentration to low concentration, driven by thermal motion. However, carrier proteins facilitate the movement of specific molecules across membranes, often against concentration gradients, through active transport or facilitated diffusion. This apparent contradiction necessitates careful experimental design to test whether carrier protein dynamics align with Fick's laws.
One approach involves single-molecule tracking techniques, such as fluorescence microscopy, to observe the movement of carrier proteins and their substrates in real-time. By labeling both the carrier protein and its substrate with fluorescent markers, researchers can track their diffusion coefficients and spatial distributions. If the substrate's movement, when bound to the carrier protein, still exhibits a concentration-dependent gradient as predicted by Fick's first law, it would suggest that the carrier protein's action does not fundamentally violate Fick's principles. However, if the substrate moves against the concentration gradient, as in active transport, Fick's laws would not directly apply.
Another experimental strategy employs fluorescence recovery after photobleaching (FRAP) to measure the mobility of carrier proteins and their substrates within membranes. In FRAP, a small region of the membrane is bleached, and the recovery of fluorescence over time is monitored. If the recovery follows a Fickian diffusion profile, characterized by a t^(1/2) dependence, it would indicate that the carrier protein's movement is consistent with Fick's second law. Deviations from this profile could suggest non-Fickian behavior, potentially due to complex interactions or energy-dependent processes.
Electrophysiological techniques, such as patch-clamp recordings, offer a complementary approach to studying carrier protein dynamics. By measuring the current flow across a membrane in response to substrate concentration changes, researchers can assess whether transport rates adhere to Fick's laws. If the current increases linearly with substrate concentration, as predicted by Fick's first law, it would support the notion that carrier proteins operate within the framework of Fickian diffusion, albeit with the assistance of protein-mediated processes.
Computational modeling also plays a vital role in testing Fick's laws in carrier protein dynamics. Molecular dynamics simulations can provide atomistic insights into the interactions between carrier proteins, substrates, and the surrounding environment. By simulating the diffusion of substrates in the presence and absence of carrier proteins, researchers can compare the resulting concentration profiles to Fickian predictions. Such models can help elucidate whether carrier proteins enhance or alter the diffusive behavior of substrates in a manner consistent with Fick's laws.
In conclusion, experimental evidence for testing Fick's laws in carrier protein dynamics requires a multifaceted approach, combining single-molecule tracking, FRAP, electrophysiology, and computational modeling. While carrier proteins introduce complexities beyond simple passive diffusion, these techniques can reveal whether their actions align with the fundamental principles of Fickian diffusion or if they operate under distinct mechanistic rules. The integration of these experimental methodologies is essential for a comprehensive understanding of carrier protein behavior in biological systems.
Statutory Law in the UK: Understanding the Basics
You may want to see also
Frequently asked questions
No, carrier proteins do not strictly obey Fick's Law. Fick's Law describes passive diffusion, where molecules move down a concentration gradient without requiring energy. Carrier proteins, however, facilitate active or facilitated transport, which may involve energy expenditure or saturation, deviating from the linear relationship predicted by Fick's Law.
The presence of carrier proteins introduces complexities that limit the applicability of Fick's Law. Carrier-mediated transport can lead to saturation at high concentrations, nonlinear flux rates, and dependence on protein density, which are not accounted for in Fick's Law's simple diffusion model.
Fick's Law is not suitable for describing transport mediated by carrier proteins. While it may approximate low-concentration behavior, it fails to capture the saturation effects, protein-specific kinetics, and potential energy requirements associated with carrier-mediated transport. Specialized models, such as the Michaelis-Menten equation, are more appropriate for such systems.