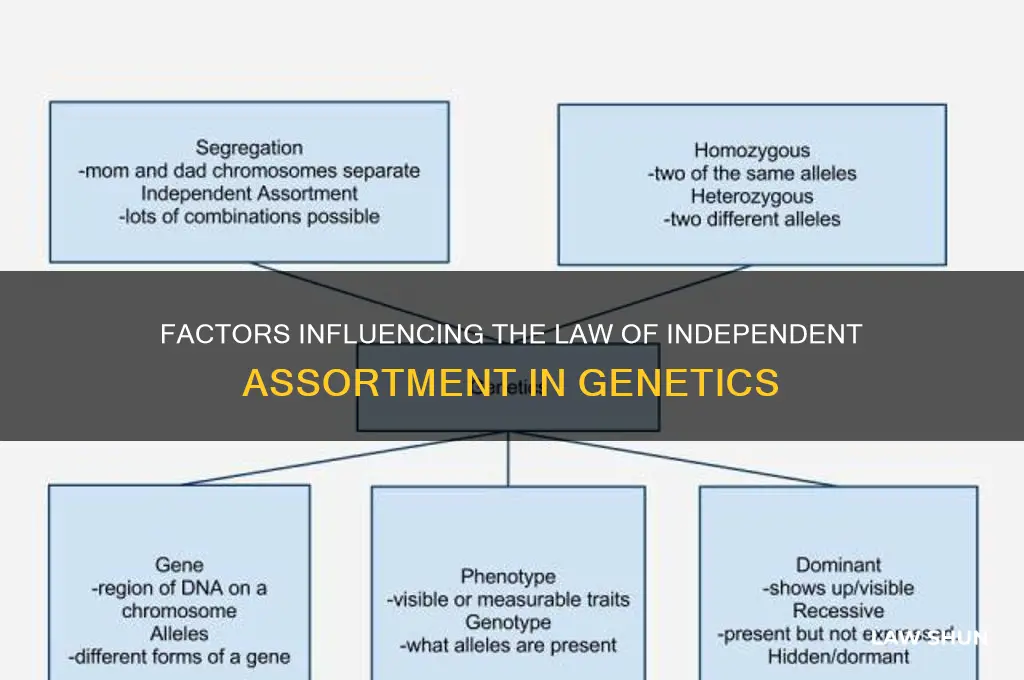
The law of independent assortment, a fundamental principle in genetics, states that alleles for different traits segregate independently during gamete formation, leading to random combinations in offspring. However, several factors can influence this law, affecting the expected 1:1:1:1 phenotypic ratio in dihybrid crosses. These factors include genetic linkage, where genes located close together on the same chromosome may not assort independently due to reduced recombination; epistasis, where the expression of one gene masks or modifies the effect of another, altering phenotypic outcomes; and environmental influences, which can impact gene expression and phenotype. Additionally, genetic interactions such as incomplete dominance, codominance, and multiple alleles can complicate the independent assortment of traits. Understanding these factors is crucial for accurately predicting genetic outcomes and interpreting inheritance patterns in complex biological systems.
| Characteristics | Values |
|---|---|
| Genetic Linkage | Closely located genes on the same chromosome assort together, violating independent assortment. |
| Chromosomal Crossovers | Frequent crossovers between homologous chromosomes can reduce linkage, restoring independent assortment. |
| Epistasis | Interaction between genes at different loci can mask independent assortment effects. |
| Polyploidy | Multiple sets of chromosomes can complicate segregation, affecting independent assortment. |
| Genetic Recombination | High recombination rates increase independent assortment by shuffling alleles. |
| Chromosomal Inversions | Inverted DNA segments reduce recombination, increasing linkage and decreasing independent assortment. |
| Genetic Distance | Greater distance between genes on a chromosome increases the likelihood of independent assortment. |
| Environmental Factors | Stressors like temperature or chemicals can influence recombination rates, indirectly affecting independent assortment. |
| Genetic Mutations | Mutations altering chromosome structure or function can disrupt independent assortment. |
| Meiosis Mechanisms | Errors in meiotic processes (e.g., synapsis, segregation) can affect independent assortment. |
Explore related products






What You'll Learn
- Genetic Linkage: Closely located genes on the same chromosome tend to be inherited together
- Crossing Over: Exchange of genetic material between homologous chromosomes during meiosis can alter assortment
- Chromosome Structure: Inversions, deletions, or duplications in chromosomes disrupt independent assortment
- Epistasis: Interactions between genes at different loci can influence phenotypic expression and assortment
- Environmental Factors: External conditions like temperature or chemicals may affect meiotic processes and gene segregation
![]()
Genetic Linkage: Closely located genes on the same chromosome tend to be inherited together
Genes located in close proximity on the same chromosome often defy the law of independent assortment, a principle that assumes genes on different chromosomes segregate independently during meiosis. This phenomenon, known as genetic linkage, means that alleles of closely situated genes are more likely to be inherited together rather than recombining freely. For instance, in fruit flies (*Drosophila melanogaster*), the genes for body color and wing shape are linked, leading to a higher frequency of co-inheritance than predicted by independent assortment. Understanding this linkage is crucial for geneticists, as it influences the predictability of trait inheritance and complicates genetic mapping efforts.
To illustrate, consider a dihybrid cross involving two linked genes. If the recombination frequency between them is low (e.g., 5%), parental combinations will dominate the offspring, deviating significantly from the 9:3:3:1 Mendelian ratio. This occurs because crossing over—the exchange of genetic material between homologous chromosomes—is less likely to separate closely located genes. However, as the distance between genes increases, the likelihood of recombination rises, gradually restoring independent assortment. Geneticists quantify this using the centimorgan (cM) unit, where 1 cM represents a 1% recombination frequency.
Practical implications of genetic linkage abound in fields like agriculture and medicine. For example, in crop breeding, linked genes for desirable traits (e.g., disease resistance and yield) can be selected together, streamlining the breeding process. Conversely, in genetic counseling, linkage can complicate risk assessments for inherited disorders. For instance, if a gene for a disorder is linked to a marker gene, the presence of the marker may indicate a higher likelihood of inheriting the disorder. Techniques like genetic mapping and linkage analysis help identify such relationships, enabling more accurate predictions.
Despite its challenges, genetic linkage also offers opportunities for scientific advancement. By studying recombination rates, researchers can construct detailed genetic maps, pinpointing the relative positions of genes on chromosomes. This is particularly valuable in organisms with complex genomes, such as humans, where linkage disequilibrium—the non-random association of alleles—provides insights into population history and disease susceptibility. For instance, the linkage between the *BRCA1* gene and certain breast cancer risk factors has informed personalized medicine approaches.
In conclusion, genetic linkage serves as a critical exception to the law of independent assortment, highlighting the intricate relationships between genes on the same chromosome. While it complicates genetic predictions, it also provides tools for understanding genome organization and function. Whether in breeding programs or medical genetics, recognizing and quantifying linkage is essential for harnessing the full potential of genetic principles. By mastering this concept, scientists can navigate the complexities of inheritance with greater precision and insight.
Legal Boundaries of Game Reviews: Understanding Rights and Responsibilities
You may want to see also
Explore related products






![]()
Crossing Over: Exchange of genetic material between homologous chromosomes during meiosis can alter assortment
During meiosis, homologous chromosomes pair up and exchange genetic material through a process called crossing over. This exchange occurs at chiasmata, physical connections between homologous chromosomes, and results in reciprocal translocation of DNA segments. While the Law of Independent Assortment predicts that alleles on different chromosomes segregate independently, crossing over introduces a wrinkle: it can link alleles on different chromosomes, altering their assortment patterns.
For example, consider two genes, one for flower color (red vs. white) and another for plant height (tall vs. short), located on different chromosomes. Normally, these traits would assort independently, resulting in a 9:3:3:1 phenotypic ratio in offspring. However, if crossing over occurs between these chromosomes, the alleles for flower color and height can become linked, leading to a deviation from the expected ratio. This phenomenon, known as genetic linkage, can be quantified using linkage maps, which measure the frequency of crossing over between genes.
To illustrate the impact of crossing over, imagine a scenario where the recombination frequency between the flower color and height genes is 20%. This means that in 20% of meioses, crossing over occurs between these genes, resulting in a reshuffling of alleles. Consequently, the observed phenotypic ratio in offspring would deviate from the expected 9:3:3:1 ratio, with an increase in the frequency of parental combinations (red-tall and white-short) and a decrease in recombinant combinations (red-short and white-tall). The extent of this deviation depends on the recombination frequency, with higher frequencies resulting in greater deviations.
When analyzing genetic data, it's essential to consider the potential effects of crossing over. One practical approach is to calculate the coefficient of coincidence (c.o.c.), which compares the observed frequency of double crossovers to the expected frequency based on single crossover rates. A c.o.c. value significantly less than 1 indicates interference, where the occurrence of one crossover inhibits the occurrence of another nearby. This phenomenon can further complicate assortment patterns, as it reduces the likelihood of multiple crossovers between linked genes. To minimize the impact of crossing over on genetic analysis, researchers can use techniques like fine-scale genetic mapping, which employs large populations and high-resolution markers to detect recombination events with greater precision.
In conclusion, crossing over serves as a critical modifier of the Law of Independent Assortment, introducing genetic linkage and altering assortment patterns. By understanding the mechanisms and consequences of crossing over, geneticists can refine their predictions, improve the accuracy of genetic mapping, and gain deeper insights into the complex interplay between genes and chromosomes. As a practical tip, when designing genetic experiments, consider the potential impact of crossing over and employ strategies to mitigate its effects, such as using larger sample sizes or selecting markers with known recombination frequencies. By doing so, researchers can unlock a more nuanced understanding of genetic inheritance and its implications for fields like genetics, evolution, and medicine.
Setting Up an LLC: Do You Need a Law License?
You may want to see also
Explore related products






![]()
Chromosome Structure: Inversions, deletions, or duplications in chromosomes disrupt independent assortment
Chromosomal abnormalities such as inversions, deletions, and duplications directly challenge the law of independent assortment by altering the physical structure and behavior of chromosomes during meiosis. Inversions, where a segment of a chromosome breaks, flips, and reattaches in the reverse orientation, can disrupt the alignment and pairing of homologous chromosomes. This misalignment often leads to uneven crossing over, resulting in gametes with unbalanced genetic material. For instance, a pericentric inversion involving the centromere can cause loops to form during meiosis I, increasing the likelihood of chromosomal breakage or mis-segregation. Such structural changes hinder the random, independent distribution of alleles, introducing predictability into genetic inheritance.
Deletions, the loss of a chromosomal segment, and duplications, the presence of extra copies of a segment, further complicate independent assortment by altering gene dosage and chromosomal pairing dynamics. A deletion on one homologous chromosome can prevent proper synapsis with its intact counterpart, leading to misalignment and non-random segregation. For example, a deletion in the *CFTR* gene region on chromosome 7 disrupts not only the gene’s function but also the orderly separation of alleles during meiosis. Duplications, on the other hand, introduce redundancy that can skew allele distribution, as seen in conditions like Charcot-Marie-Tooth disease, where duplicated genes on chromosome 17 interfere with normal meiotic processes. These structural anomalies force certain alleles to co-segregate, violating the principle of independent assortment.
To illustrate the practical implications, consider the case of chromosomal inversions in *Drosophila melanogaster*. Inverted chromosome regions in this model organism often result in reduced fertility due to the formation of unbalanced gametes. For researchers studying genetic inheritance, identifying such inversions is critical, as they can be detected through cytogenetic techniques like Giemsa banding or molecular methods such as comparative genomic hybridization (CGH). Similarly, deletions and duplications in humans, often identified through karyotyping or microarray analysis, require careful genetic counseling, as they can lead to conditions like Cri-du-chat syndrome (deletion on chromosome 5) or Down syndrome (trisomy 21). Understanding these structural disruptions is essential for predicting inheritance patterns and managing genetic disorders.
While inversions, deletions, and duplications disrupt independent assortment, they also highlight the dynamic nature of chromosomal evolution. Inversions, for instance, can act as evolutionary drivers by suppressing recombination in heterozygotes, allowing beneficial gene combinations to persist. However, this comes at the cost of reduced genetic diversity in affected regions. Deletions and duplications, though often deleterious, contribute to genomic plasticity, enabling species to adapt to changing environments over time. For geneticists and clinicians, recognizing these dual roles—as disruptors of Mendelian principles and as agents of evolutionary change—is crucial for both theoretical understanding and practical applications in genetics.
In summary, chromosomal inversions, deletions, and duplications serve as potent disruptors of the law of independent assortment by altering chromosome structure and behavior during meiosis. These abnormalities introduce predictability into allele segregation, often leading to unbalanced gametes and genetic disorders. Yet, they also underscore the complexity of genomic evolution, offering insights into both the challenges and opportunities presented by chromosomal variability. For those navigating the intricacies of genetics, whether in research or clinical settings, understanding these structural disruptions is indispensable for interpreting inheritance patterns and addressing their consequences.
Understanding Legal Phrases: Do Years of Laws Matter for You?
You may want to see also
Explore related products




![Independent [Stage-10] 215mm Silver Skateboard Trucks (Set Of 2)](https://m.media-amazon.com/images/I/515vEhiVQJL._AC_UL320_.jpg)

![]()
Epistasis: Interactions between genes at different loci can influence phenotypic expression and assortment
Genes rarely act in isolation; their interactions can profoundly alter phenotypic outcomes, challenging the simplicity of the law of independent assortment. Epistasis, where genes at different loci influence each other’s expression, serves as a prime example of this complexity. Consider coat color in mice: the *Agouti* gene determines pigment distribution, but its effects are suppressed by a dominant allele at the *Extension* locus, resulting in a uniform yellow coat. Here, the *Extension* gene epistatically masks the *Agouti* gene’s phenotype, demonstrating how independent assortment can be disrupted by inter-locus interactions.
To understand epistasis, think of genes as players in a biological orchestra, where one musician’s performance can silence or amplify another’s. For instance, in humans, the *HBB* gene encodes beta-globin, essential for hemoglobin function. However, mutations in the *BCL11A* gene, located on a different chromosome, can epistatically enhance fetal hemoglobin production, offering a therapeutic target for sickle cell anemia. This interaction highlights how genes at distinct loci can modulate phenotypic expression, bypassing the expected independence of assortment.
Practical implications of epistasis extend to agriculture and medicine. In crop breeding, epistatic interactions between genes controlling yield and disease resistance can limit the predictability of trait inheritance. For example, a gene conferring drought tolerance might only express its phenotype when paired with a specific allele at another locus. Breeders must account for these interactions to optimize outcomes. Similarly, in pharmacogenomics, epistasis can influence drug efficacy; a patient’s response to a medication may depend on the combined effects of variants at multiple loci, not just one.
To navigate epistasis, researchers employ quantitative genetics and bioinformatics tools. Genome-wide association studies (GWAS) can identify epistatic interactions by analyzing how allele combinations at different loci correlate with phenotypes. For instance, a GWAS on hypertension might reveal that the combined presence of variants in the *AGT* and *ACE* genes significantly elevates blood pressure, even if each variant alone has a minor effect. Such analyses underscore the need to move beyond single-gene models when studying complex traits.
In conclusion, epistasis serves as a reminder that genetic inheritance is not always a straightforward sum of independent parts. By recognizing how genes at different loci interact, scientists can better predict phenotypic outcomes, improve breeding strategies, and personalize medical treatments. Whether in a lab, clinic, or field, understanding epistasis is essential for unraveling the intricacies of genetic assortment and its real-world applications.
When Courts Strike Down Laws: Real-Life Unconstitutional Declarations Explained
You may want to see also
Explore related products






![]()
Environmental Factors: External conditions like temperature or chemicals may affect meiotic processes and gene segregation
External conditions, such as temperature fluctuations and chemical exposures, can subtly yet significantly disrupt the delicate processes of meiosis, thereby influencing gene segregation and challenging the predictability of the law of independent assortment. For instance, elevated temperatures during meiosis in *Drosophila melanogaster* have been shown to increase chromosomal missegregation, leading to higher rates of aneuploidy. This occurs because heat stress can impair the function of spindle assembly checkpoint proteins, which are critical for accurate chromosome alignment and segregation. Similarly, in plants like *Arabidopsis thaliana*, exposure to temperatures exceeding 28°C during meiosis results in reduced synapsis efficiency and increased crossover interference, altering the independent assortment of alleles. These examples underscore how temperature acts as a potent environmental modifier of genetic outcomes.
Chemical exposures provide another layer of complexity, with specific compounds directly targeting meiotic machinery. For example, the fungicide prothioconazole, commonly used in agriculture, disrupts microtubule dynamics in mammalian oocytes at concentrations as low as 10 μM. This interference leads to misaligned chromosomes and non-disjunction events, skewing the expected 1:1 segregation ratio. In yeast, exposure to the alkylating agent methyl methanesulfonate (MMS) induces DNA damage during meiosis, triggering error-prone repair mechanisms that can shuffle genetic material unpredictably. Such chemical-induced alterations highlight the vulnerability of meiotic processes to external agents, particularly in occupational or agricultural settings where exposure is chronic.
To mitigate these effects, researchers and practitioners must adopt proactive strategies. For laboratory studies, maintaining tight temperature control (±0.5°C) during meiotic experiments is essential, especially for organisms like *Caenorhabditis elegans*, which exhibit temperature-sensitive meiotic defects above 25°C. In agricultural contexts, rotating fungicides with different modes of action can reduce the risk of prothioconazole-induced meiotic errors in crops. Additionally, monitoring environmental chemical levels, particularly in regions with heavy pesticide use, can help identify at-risk populations. For instance, communities near vineyards in California have reported higher incidences of chromosomal abnormalities in newborns, correlating with increased prothioconazole usage.
Comparatively, while genetic factors like gene mutations are intrinsic and immutable, environmental factors offer a degree of control and prevention. Unlike inherited defects in meiotic proteins, temperature and chemical exposures can be regulated through behavioral and policy interventions. For example, the European Union’s restriction of prothioconazole use in residential areas demonstrates how regulatory measures can safeguard genetic integrity. This contrasts with the challenges of addressing genetic predispositions, which require complex gene therapies or selective breeding programs. By focusing on environmental modifiers, we can achieve more immediate and scalable solutions to preserve the fidelity of independent assortment.
In conclusion, environmental factors like temperature and chemicals serve as dynamic modulators of meiotic processes, introducing variability into gene segregation that defies the simplicity of the law of independent assortment. From heat-induced chromosomal misalignment in fruit flies to chemical-driven DNA damage in yeast, these external conditions highlight the fragility of meiotic mechanisms. Practical steps, such as temperature regulation and chemical exposure monitoring, offer tangible ways to mitigate these effects. By recognizing the interplay between environment and genetics, we can better predict and control the outcomes of inheritance, ensuring the stability of genetic systems in both natural and managed contexts.
Is Nigeria an Islamic Country by Law? Exploring Its Legal Framework
You may want to see also
Frequently asked questions
The Law of Independent Assortment, proposed by Gregor Mendel, states that alleles for different traits segregate independently during the formation of gametes, provided the genes are located on different chromosomes or are far enough apart on the same chromosome.
Gene linkage occurs when genes are located close together on the same chromosome, causing them to be inherited together more often than predicted by independent assortment. This reduces the independent segregation of alleles, leading to deviations from the expected ratios.
Yes, crossing over during meiosis can influence the Law of Independent Assortment. Crossing over between homologous chromosomes can shuffle alleles, allowing linked genes to assort independently if the crossover occurs between them.
The Law of Independent Assortment applies to genes on different chromosomes. In organisms with more chromosomes, there are more opportunities for independent assortment, as each chromosome pairs and segregates independently during meiosis.
Epistasis, where one gene masks or modifies the expression of another, does not directly affect the Law of Independent Assortment. However, it can complicate the observed phenotypic ratios, making it seem like genes are not assorting independently, even if they are.